
Cafergot
"Cheap cafergot 100 mg without a prescription, pain treatment center houston".
By: O. Ugolf, M.B. B.CH. B.A.O., Ph.D.
Associate Professor, George Washington University Medical School
Molecular complexity and its impact on the probability of finding leads for drug discovery neck pain treatment kerala discount cafergot online amex. The genesis of high-throughput structure-based drug discovery using protein crystallography pain tmj treatment buy cafergot in india. Making drugs on proteins: sitedirected ligand discovery for fragment-based lead assembly myofascial pain treatment uk order cafergot 100 mg without prescription. Algorithms for the automated selection of fragment-like molecules using singlepoint surface plasmon resonance measurements heel pain treatment urdu purchase cafergot from india. Mode of interaction of Я-hydroxy-Яmethylglutaryl coenzyme A reductase with strong binding inhibitors: compactin and related compounds. Stromelysin inhibitors designed from weakly bound fragments: effects of linking and cooperativity. Structure of Bcl-xL-Bak peptide complex: recognition between regulators of apoptosis. De novo fragment design: a medicinal chemistry approach to fragment-based lead generation. Methods and Principles in Medicinal Chemistry 2007, 36(Mass Spectrometry in Medicinal Chemistry), 321338. Substrate activity screening: a fragment-based method for the rapid identification of nonpeptidic protease inhibitors. Fragment-based substrate activity screening method for the identification of potent inhibitors of the Mycobacterium tuberculosis phosphatase PtpB. Click chemistry in situ: acetylcholinesterase as a reaction vessel for the selective assembly of a femtomolar inhibitor from an array of building blocks. In situ selection of lead compounds by click chemistry: target-guided optimization of acetylcholinesterase inhibitors. Virtual fragment screening for novel inhibitors of 6-phosphogluconate dehydrogenase. Fragment-based discovery of mexiletine derivatives of bioavailable inhibitors of urokinase-type plasminogen activator. Role of plasmin and gelatinase in extracellular matrix degradation by cultured rat mesangial cells. Design of novel and selective inhibitors of urokinase-type of plasminogen activator with improved pharmacokinetic properties for use as antimetastatic agents. Ьber eine neue Klasse von Verbindungen mit spasmolytischer und zentral analgetischer Wirksamkeit unter besonderer Berьcksichtigung des 1-Methyl-4-phenyl-piperidin-4-дure-дthylesters (Dolantin). Alcohols of the 6,14-endo-ethenotetrahydrooripavine series and derived analogs of N-allylnormorphine and norcodeine. New diarylmethylpiperazines as potent and selective nonpeptidic х opiod receptor agonists with increased in vitro metabolic stability. The Sulfonamides and Allied Compounds, American Chemical Society Monograph Series, Reinhold: New York, 1948. A common pharmacophore for taxol and the epothilones based on the biological activity of a taxane molecule lacking a C-13 side chain. Kinetics and thermodynamics of binding of fluorescent taxol derivatives to an exposed site. Intravenous ethanol for the treatment of alcohol withdrawal syndrome in critically ill patients. The identification of toxicophores for the prediction of mutagenicity, hepatotoxicity and cardiotoxicity. Structural alert/reactive metabolite concept as applied in medicinal chemistry to mitigate the risk of idiosyncratic drug toxicity: a perspective based on the critical examination of trends in the top 200 drugs marketed in the United States. Korolkovas, A Essentials of Molecular Pharmacology: Background for Drug Design; Wiley: New York, 1970, pp 5457. In silico techniques for the identification of bioisosteric replacements for drug design. A review on bioisosterism: A rational approach for drug design and molecular modification.
These differences in reflectivity can be explained either by variations in the packing fraction of the regolith or by variations in its depth joint and pain treatment center santa maria ca purchase cafergot online now. The latter explanation appears more physically realistic if one also takes into account the results of Earth-based observations pain treatment satisfaction questionnaire purchase cafergot 100mg with mastercard. The Apollo 14 and 15 missions provided the opportunity to continue bistatic radar investigations at wavelengths of 13 cm and 116 cm (Tyler and Howard treating pain for uti best purchase cafergot, 1973) pain treatment for rheumatoid arthritis buy cafergot with a visa. The signal-to-noise ratio and the surface resolution of these measurements were both an order of magnitude better than previous experiments of the same type. Although the extent of the datasets obtained was limited, the investigators found regions of the lunar surface whose physical properties could not easily be explained by simple models. For = 13 cm, mare surfaces could be modeled well using a value of the dielectric constant = 3. At = 116 cm, much of the mare could also be modeled simply with a similar dielectric constant, but in this case, considerable deviations from the simple characterization were observed. No successful models for the structure and properties of the anomalous regions could be developed; the authors speculated that their data implied the existence of some frequency dispersive mechanism, such as layering in a high-loss dielectric medium. In highland regions the longer wavelength (116 cm) data were fitted successfully to a model using a uniform dielectric interface with = 2. Observations at the shorter wavelength (13 cm) in the Apennine Mountains and in the central highland regions also yielded consistent results, but the data from units in the Palus Somni area and the rim of Mare Crisium could not be fitted with simple models at = 13 cm. Since the polarization of returned radio signals can be determined, the echoes from the lunar surface also contain information about slopes and surface roughness. The complexity of both the surface and the physical mechanisms that alter the polarization of the signal permit only some general statements with any degree of confidence. At = 13 cm, highland rms (root-mean-square) slopes range from 6° to 8°, while mare rms slopes range from 2° to 4° (Tyler and Howard, 1973). Although values derived from the 116-cm observations agree with the 13-cm data in the highlands, the mare values are only half as large as those derived from the 13-cm data. Little work has yet been done on the lunar surface itself to verify the surface structures inferred from radar scattering behavior. The very low electrical loss characteristics of the lunar regolith permitted detection of echoes from subsurface features as well as from the surface itself. First, the relatively weak Physical Properties of the Lunar Surface 565 signals returned from subsurface reflectors are obscured by strong surface clutter. Second, time delays associated with signals from subsurface features cannot be unambiguously distinguished from those associated with surface features lying some distance from the ground track. These problems have been overcome to some extent in the data from flat mare regions by correlating reflections from adjacent ground tracks. This correlation allows discrimination between subsurface layers and long linear surface features, which coincidentally parallel the orbital path and produce reflections at a distance from it (Peeples et al. Thus far, reflecting subsurface layers have been detected with confidence in both Mare Serenitatis and Mare Crisium. The placement and extent of the reflectors beneath Mare Serenitatis have been used to infer the geological history of the basin (see. These profiles, which extended along nearly two complete lunar orbits, were constructed from sampling at intervals of about. Temperature profiles across Aristarchus Crater, a large central-peak crater emplaced in a mare region. Profiles originate at the center of the crater and traverse the crater in the directions indicated (southwest, south-southwest, and south-southeast). The higher pre-sunrise temperatures of the center indicate that much of the crater interior consists of large blocks of exposed bedrock. The nighttime temperatures fall significantly along the crater floor and are low but somewhat variable across the skirt of continuous ejecta around the crater. The ejecta deposits that occur beyond the limit of near-crater continuous ejecta and as far out as three crater radii (60 km) from the rim have smooth and uniform temperature profiles, unlike the irregular mare surface that is exposed beyond the ejecta. They also published very accurate (better than ±25 m in elevation) profiles of Mare Crisium and Mare Serenitatis. This instrument collected data on lunar surface emissions in the wavelength range from 1.
Buy 100 mg cafergot otc. (How To Get Rid of Abdominal Scar Tissue with Jill Miller).
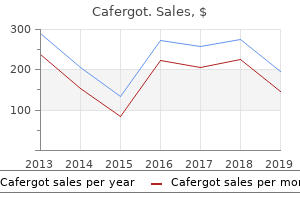
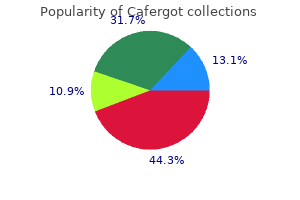
Once glycogen stores have been depleted mtus chronic pain treatment guidelines generic cafergot 100 mg with amex, the replenishment of these stores is more prolonged in horses than in other species taking on average 72 hours following glycogen depleting exercise pain treatment in multiple sclerosis buy cafergot in india. One study found that horses fed a diet high in non-structural carbohydrates compared to a diet higher in fat had quicker glycogen replenishment (Veronique et al pain treatment options discount cafergot online mastercard. Specifically pain treatment center bismarck nd cafergot 100mg sale, the density of mitochondria will increase with training, leading to an increased capacity for oxidative phosphorylation. Diet and physical conditioning of the animal play a major role in the utilization of energy substrates. A mix of both carbohydrates and fat is metabolized to produce energy during work, and fatigue is a factor of a depletion in muscle glycogen, reduction in circulating glucose, and a decrease in intracellular pH related to an increased production of lactate. Digestible energy values for feedstuffs are typically estimates based on the proximate analysis of other components of the diet (digestible protein, crude fiber, nitrogen free extract, and ether extract). A score of 1 indicates a highly starved animal while a score of 9 coincides with a morbidly obese individual. Dietary carbohydrates can be classified into two general categories: non-structural and structural carbohydrates. Exceeding this limit may result in starch leaking into the hindgut of the horse resulting in digestive upset. It is always recommended to reduce large volume feedings in to smaller feedings multiple times per day. Feeding fat allows for the development of a more energy dense diet and incoroporation of certain fatty acids that are vital for cell membranes, skin and coat quality, and hoof health. Fat sources are variable, and the source of the fat dictates the fatty acid profile. Dietary fat also facilitates the absorption of fat-soluble vitamins that are important to numerous physiological processes. The upper limit of fat inclusion to the diet should be no greater than 20-25% of the total diet (not the individual components). Considering the type of exercise the horse is performing is important in determining the fueling strategies. During high bouts of intense exercise, almost all the energy is derived from creatine phosphate, glucose, and glycogen via anaerobic respiration (Hinchcliff et al. Conversely, longer and less intense exercise bouts can be fueled by a diet rich in fats. Protein and the Exercising Horse As the exercise level of the horse increases as dictated by Table 1, the dietary protein requirements increase as well. However, these needs are typically met as the dry matter intake increases to meet the elevated energy requirements. While protein content is important, protein quality is essential to the functionality of the performance horse. The adage that "not all protein is created equally" holds true for the performance horse diet. While most amino acid requirements for horses are not fully elucidated, there is an established lysine requirement in horses. Typically, commercially available concentrate feeds are fortified appropriately for the classes of horses they are designed to feed relative to the amino acid balance. High quality protein sources for horses include but are not limited to soybean meal, alfalfa, and whey protein, in addition to individual crystalline amino acids. Performance horses often are fed diets with excess dietary protein due to the goal of meeting the increased energy requirements of the horse. The horse is fairly tolerant of excess protein in the diet with excess amino acids being broken down to nitrogen and ammonia and being excreted in the urine and feces. If adequate clean water is not available, the horse can be at risk for dehydration or kidney dysfunction. Further, excreted ammonia can be an irritant to mucus membranes and may aggravate the respiratory tract. Water and the Performance Horse Water is often the most overlooked nutrient regardless of the class of horse for which a diet is being formulated.
Ai Ai -d Bi pain treatment research discount cafergot 100 mg, - d k degradation with reaction rate kd knee pain treatment by physiotherapy purchase cafergot 100mg without prescription, the mean life time of a 1 molecule can be expressed as r = kd pain medication for a uti purchase cafergot pills in toronto. The diffusion can also be considered as a first order reaction with the reaction rate constant d = D/l2 pain treatment pregnancy buy cafergot on line amex. Hence, the mean free time for a molecule staying within a bin can be expressed as d = l2 /D. Here we will derive a more specified expression for the ideal discretization length. Apparently, the smaller the voxel size is, the more accurate the resulted system will be. However, a small voxel size often results in large propensities for species jumping to neighboring voxels, but the heavy computational cost on the back-andforth jumping makes little contribution to the actual population distribution evolution. Thus, it is important to find a optimal voxel size that is as large as possible while still maintaining a reasonable simulation error. The ideal discretization should be based on the assumption that molecules of a species within a voxel are well-stirred, such that any two copies of that species within a voxel have similar probability distributions before one of them fires a reaction. By the finite-difference schema, the propensity function for the reaction diffusion system (6) can be formulated as kd Ai, (7) Ai+1 - 2Ai + Ai-1 a2 (Ai) = D, l2 where Ai denotes the population of A at the i-th bin and l is the length of the bin. The assumption of the discretization requires that the lengths of the bins be small enough, such that species within the bins can be considered well stirred. Previous work [12] has shown that this criterion is equivalent to r 1, (8) d where r is the mean free time with respect to the reactive collision and d denotes the mean free time, during which a molecule will remain within a bin. For the first order a1 (Ai) = where (x) is the Dirac delta function, with - (x)dx = 1. The solutions to the two diffusion functions are u1 (x, t) u2 (x, t) = = 1 e- 4Dt, 4Dt (x-l)2 1 e- 4Dt 4Dt x2. In probability theory, the Kullback-Leibler divergence is a non-symmetric measure of the difference between two probability distribution functions. In the simple reaction diffusion model, the mean life time of reaction is r = 1/kd. If we set this threshold as 5%, we will have an analytic solution for the critical discretization size: 2 lc = 0. The critical discretization size with respect to the conservative threshold is: 2 lc = 0. We can see from the plot figure 1 that, When the discretization size reduces to 0. In this simple model, the 5% K-L divengence threshold is good enough to obtain an accurate simulation. Theoretical Analysis Typical biological systems contain multiple species and reactions. From equation (14) we can see that fast reaction (large kd) and slow diffusion (small D) lead to small discretization size l. For a multispecies system, if a uniform discretization is used, l has to using the largest kd and the smallest D, and a small discretization size l leads to heavy computational burden. Here with the ideal discretization size calculated in previous section, we propose a multiscale discretization method, which assigns a proper discretization size to each species, if necessary, depending on the diffusion rates and the chemical kinetics. The domain is partitioned differently for different species, according to the diffusion and reaction rate constants of each species. Each species in the domain is represented by the state vector Xi (t) = [Xi,1 (t). For a larger l, it will lead to larger simulation error with respect to the homogeneous assumption. However, the optimal discretization size for this species still depends on the time scales of diffusion and reaction. Equation (14) and (15) can still be applied, although kd will be the overall change rate of this species rather than the reaction rate for a simple reaction. Numerical Experiment Below we present the numerical experiment results for the simple reaction-diffusion model (6).